ペプチドは、ペプチド結合を介した複数のアミノ酸の接続によって形成される化合物のクラスです。それらは生きている生物で遍在しています。これまで、生物には数万個のペプチドが発見されています。ペプチドは、さまざまなシステム、臓器、組織、細胞、および生活活動の機能的活動の調節に重要な役割を果たし、機能分析、抗体研究、医薬品開発、その他の分野でよく使用されます。バイオテクノロジーとペプチド合成技術の開発により、ますます多くのペプチド薬が開発され、診療所で適用されています。
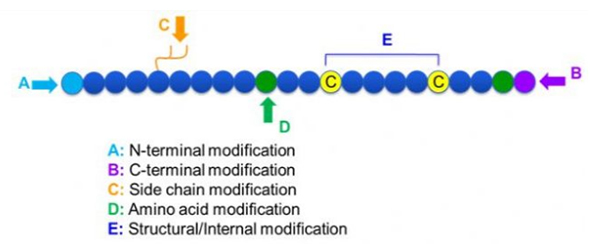
多種多様なペプチド修飾があります。これは、ポスト修飾とプロセス修飾(導出されたアミノ酸修飾を使用)、およびN末端修飾、C末端修飾、サイドチェーン修飾、アミノ酸修飾、スケルトン修飾などに単純に分割できます(図1)。ペプチド鎖の主要な鎖構造またはサイドチェーングループを変化させる重要な手段として、ペプチド修飾はペプチド化合物の物理的および化学的特性を効果的に変化させ、水溶解度を高め、in vivoでの作用時間を延長し、免疫力を排除し、毒性副作用を減らし、毒性副作用を減らします。
1。環化
環状ペプチドには生物医学に多くの用途があり、生物活性を持つ多くの天然ペプチドは環状ペプチドです。環状ペプチドは線形ペプチドよりも剛性がある傾向があるため、消化器系に非常に耐性があり、消化管で生存し、標的受容体に対してより強い親和性を示します。環化は、特に大きな構造的骨格を持つペプチドにとって、環状ペプチドを合成する最も直接的な方法です。環化モードに応じて、サイドチェーンサイドチェーンタイプ、端子 - サイドチェーンタイプ、端子 - 端子タイプ(端から端まで)に分割できます。
(1)Sidechain-to-Sidechain
側鎖環化への最も一般的なタイプの側鎖環化は、システイン残基間のジスルフィド橋渡しです。この環化は、脱保護され、酸化されてジスルフィド結合を形成するために酸化されるシステイン残基のペアによって導入されます。ポリクリック合成は、スルフヒドリル保護群を選択的に除去することにより実現できます。環化は、浸透後の溶媒または前処理樹脂のいずれかで行うことができます。樹脂の環化は、樹脂のペプチドが環状構造を容易に形成しないため、溶媒環化よりも効果が低い場合があります。別のタイプの側鎖 - 側鎖環化環化は、アスパラギン酸またはグルタミン酸残基と塩基アミノ酸との間のアミド構造の形成です。これには、サイドチェーン保護グループを樹脂または解離後のポリペプチドから選択的に除去する必要があります。サイドチェーンの3番目のタイプ - サイドチェーン環化は、チロシンまたはp-ヒドロキシフェニルグリシンによるジフェニルエーテルの形成です。天然産物におけるこのタイプの環化は微生物製品にのみ見られ、環化製品はしばしば潜在的な薬用価値を持っています。これらの化合物の調製には、独自の反応条件が必要なため、従来のペプチドの合成にはあまり使用されません。

(2)ターミナルからサイドシェイン
端子側鎖環化は通常、リジンまたはオルニチン側鎖のアミノ基を伴うC末端、またはアスパラギン酸またはグルタミン酸側鎖を備えたN末端を含みます。他のポリペプチド環化は、末端Cとセリンまたはスレオニン側鎖の間にエーテル結合を形成することにより作られています。
(3)ターミナルまたはヘッドツーテールタイプ
鎖のポリペプチドは、溶媒で循環するか、サイドチェーンサイクルによって樹脂で固定することができます。ペプチドのオリゴマー化を避けるために、溶媒集中化に低濃度のペプチドを使用する必要があります。頭から尾の合成リングポリペプチドの収率は、鎖ポリペプチドの配列に依存します。したがって、環状ペプチドを大規模に調製する前に、最初に鎖鎖ペプチドのライブラリーを作成し、次に環化して最良の結果を伴う配列を見つける必要があります。
2。N-メチル化
N-メチル化はもともと天然のペプチドで発生し、水素結合の形成を防ぐためにペプチド合成に導入され、それにより生分解とクリアランスによりペプチドがより耐性になります。 N-メチル化アミノ酸誘導体を使用したペプチドの合成が最も重要な方法です。さらに、N-(2-ニトロベンゼンスルホニル塩化物)ポリペプチド - レシン中間体とメタノールとのミツノブ反応も使用できます。この方法は、N-メチル化アミノ酸を含む環状ペプチドライブラリーの調製に使用されています。
3。リン酸化
リン酸化は、自然界で最も一般的な翻訳後修飾の1つです。ヒト細胞では、タンパク質の30%以上がリン酸化されています。リン酸化、特に可逆的なリン酸化は、シグナル伝達、遺伝子発現、細胞周期および細胞骨格の調節、アポトーシスなどの多くの細胞プロセスを制御する上で重要な役割を果たします。
リン酸化はさまざまなアミノ酸残基で観察できますが、最も一般的なリン酸化標的はセリン、スレオニン、およびチロシン残基です。ホスホチロシン、ホスホトレオニン、およびホスホセリン誘導体は、合成中にペプチドに導入するか、ペプチド合成後に形成されることができます。選択的リン酸化は、保護基を選択的に除去するセリン、スレオニン、チロシンの残基を使用して達成できます。一部のリン酸化試薬は、後修飾によりリン酸基をポリペプチドに導入することもできます。近年、リジンの部位特異的リン酸化は、化学的に選択的なシュッディンガー - リン酸塩反応を使用して達成されています(図3)。

4。ミリストイル化とパルミトイル化
N末端の脂肪酸をアシル化すると、ペプチドまたはタンパク質が細胞膜に結合することができます。 N末端のミリダモイル化された配列により、SRCファミリープロテインキナーゼと逆転写酵素GAQタンパク質が細胞膜に結合することを標的とすることができます。ミリスチン酸は、標準的な結合反応を使用して樹脂 - ポリペプチドのN末端に関連しており、得られるリポペプチドは標準条件下で解離し、RP-HPLCによって精製される可能性があります。
5。グリコシル化
バンコマイシンやテイコラニンなどのグリコペプチドは、薬物耐性細菌感染症の治療のために重要な抗生物質であり、他のグリコペプチドは免疫系を刺激するためにしばしば使用されます。さらに、多くの微生物抗原はグリコシル化されているため、感染の治療効果を改善するためにグリコペプチドを研究することは非常に重要です。一方、腫瘍細胞の細胞膜上のタンパク質は異常なグリコシル化を示すことがわかっています。グリコペプチドは、FMOC/T-BU法によって調製されます。スレオニンやセリンなどのグリコシル化残基は、グリコシル化アミノ酸を保護するために、ペンタフルオロフェノールエステル活性化FMOCによってポリペプチドにしばしば導入されます。
6。イソプレン
イソペンタジエニル化は、C末端近くの側鎖のシステイン残基で発生します。タンパク質イソプレンは、細胞膜親和性を改善し、タンパク質間相互作用を形成することができます。イソペンタジエン化タンパク質には、チロシンホスファターゼ、小さなGTase、コチャペロン分子、核層、およびセントロメア結合タンパク質が含まれます。イソプレンポリペプチドは、樹脂上のイソプレンを使用して、またはシステイン誘導体を導入することにより調製できます。
7。ポリエチレングリコール(PEG)修飾
PEGの修飾は、タンパク質の加水分解安定性、生体内分布、ペプチド溶解度を改善するために使用できます。ペプチドへのPEG鎖の導入は、その薬理学的特性を改善し、タンパク質分解酵素によるペプチドの加水分解を阻害する可能性があります。 PEGペプチドは、糸球体毛細血管断面積を通常のペプチドよりも簡単に通過し、腎クリアランスを大幅に減らします。 in vivoでのPEGペプチドの活性半減期が拡張されているため、通常の治療レベルは、より低い用量と頻度の低いペプチド薬で維持できます。ただし、PEGの変更にも悪影響があります。大量のPEGは、酵素がペプチドを分解するのを防ぎ、標的受容体へのペプチドの結合も減少させません。しかし、PEGペプチドの低親和性は通常、より長い薬物動態の半減期によって相殺され、体内に長く存在することにより、PEGペプチドは標的組織に吸収される可能性が高くなります。したがって、PEGポリマーの仕様は、最適な結果のために最適化する必要があります。一方、PEGペプチドは、腎クリアランスの減少により肝臓に蓄積し、高分子症候群を引き起こします。したがって、ペプチドを薬物検査に使用する場合、PEGの修正をより慎重に設計する必要があります。

PEG修飾子の一般的な修飾グループは、アミノ(-amine)-NH2、アミノメチル-CH2 -NH2、ヒドロキシ-OH、カルボキシ-COOH、スルフヒドリル( - チオール)-SH、マレイミド - マルス、マリド - マル、サッカー酸アセッツ酸アセッツ酸アセッツ酸アセッツ酸塩-SCM、サッサイミドアセッツ酸塩-SC、サッサミドアセッツ酸塩-SC、 n-ヒドロキシシシニミド-NHS、アクリレート-CH2CH2COOH、アルデヒド-CHO(プロピオン - ナルド、ブチラルドなど)、アクリル塩基(-acrylate-Acrl)、アジド - アジド、ビオチニル - ビオチン、フルオレシン、グタリル-GA、アキラジドアルキン - アルキン、p-トルエンスルホン酸-OTS、コキシミドコハク酸-SSなど。カルボン酸を伴うPEG誘導体は、N末端アミンまたはリジン側鎖に結合することができます。アミノ活性化PEGは、アスパラギン酸またはグルタミン酸側鎖に結合できます。 MAL活性化PEGは、完全に保護されたシステイン側鎖のメルカプタンに共役することができます[11]。 PEG修飾子は一般に次のように分類されます(注:MPEGはメトキシ-PEG、CH3O-(CH2CH2O)N-CH2CH2-OH):
(1)ストレートチェーンペグ修飾子
MPEG-SC、MPEG-SCM、MPEG-SPA、MPEG-OTS、MPEG-SH、MPEG-ALD、MPEG-BUTYRALD、MPEG-SS
(2)二機能性PEG修飾子
HCOO-PEG-COOH、NH2-PEG-NH2、OH-PEG-COOH、OH-PEG-NH2、HCL・NH2-PEG-COOH、MAL-PEG-NHS
(3)分岐PEG修飾子
(mpeg)2-nhs、(mpeg)2-ald、(mpeg)2-nh2、(mpeg)2 mal
8。生物化
ビオチンはアビジンまたはストレプトアビジンと強く結合することができ、結合強度は共有結合にさえ近い。ビオチン標識ペプチドは、免疫測定法、組織細胞化学、および蛍光ベースのフローサイトメトリーで一般的に使用されています。標識された抗ビオチン抗体は、ビオチン化ペプチドに結合するためにも使用できます。ビオチンラベルは、多くの場合、リジン側鎖またはN端子に取り付けられています。 6-アミノカプロ酸は、ペプチドとビオチンの間の結合としてよく使用されます。結合は基質への結合に柔軟であり、立体障害の存在下でよりよく結合します。
9。蛍光標識
蛍光標識は、生細胞のポリペプチドを追跡し、酵素と作用機構を研究するために使用できます。トリプトファン(TRP)は蛍光であるため、固有の標識に使用できます。トリプトファンの放出スペクトルは、末梢環境に依存し、ペプチド構造と受容体結合の検出に役立つ溶媒極性の減少とともに減少します。トリプトファン蛍光は、プロトン化アスパラギン酸とグルタミン酸によって消すことができ、その使用を制限する可能性があります。ダンシル塩化物基(ダンシル)は、アミノ基に結合すると非常に蛍光があり、アミノ酸またはタンパク質の蛍光標識としてよく使用されます。
蛍光共鳴エネルギー変換(FRET)は、酵素研究に役立ちます。 FRETを適用すると、基質ポリペプチドには通常、蛍光標識基と蛍光消されるグループが含まれています。標識された蛍光基は、非光子エネルギー伝達を通じてクエンチャーによってクエンチされます。問題の酵素からペプチドが解離すると、標識群は蛍光を放出します。
10。ケージポリペプチド
ケージペプチドには、ペプチドが受容体への結合から保護する光学的に取り外し可能な保護基があります。紫外線にさらされると、ペプチドが活性化され、受容体への親和性が回復します。この光学的活性化は、時間、振幅、または位置に従って制御できるため、ケージペプチドを使用して細胞で発生する反応を研究できます。ケージポリペプチドで最も一般的に使用される保護基は、2-ニトロベンジル基とその誘導体であり、保護アミノ酸誘導体を介してペプチド合成で導入できます。開発されたアミノ酸誘導体は、リジン、システイン、セリン、チロシンです。ただし、アスパラギン酸およびグルタミン酸誘導体は、ペプチド合成と解離中の環化に対する感受性のため、一般的には使用されていません。
11。多形成ペプチド(MAP)
通常、短いペプチドは免疫がなく、抗体を産生するためにキャリアタンパク質に結合する必要があります。多形成ペプチド(MAP)は、リジン核に接続された複数の同一のペプチドで構成されています。これは、高効力免疫原性を特異的に発現させることができ、ペプチドキャリアタンパク質coupleteの調製に使用できます。 MAPポリペプチドは、MAP樹脂の固相合成により合成できます。ただし、不完全な結合により、一部の枝にペプチド鎖が欠落または切り捨てられた場合、元のMAPポリペプチドの特性は示されません。別の方法として、ペプチドを個別に調製および精製し、マッピングするために結合することができます。ペプチドコアに取り付けられたペプチド配列は明確に定義されており、質量分析によって簡単に特徴付けられます。
結論
ペプチド修飾は、ペプチドを設計する重要な手段です。化学的に修飾されたペプチドは、高い生物学的活性を維持するだけでなく、免疫原性と毒性の欠点を効果的に回避することもできます。同時に、化学修飾は、いくつかの新しい優れた特性をペプチドに導くことができます。近年、ポリペプチドの修正後のC-H活性化の方法が急速に開発されており、多くの重要な結果が達成されています。
投稿時間:2025-07-03